Is there any chance of predicting the history of life? On the footsteps of Gould, that would seem a foolish quest. Rewind the tape and replay it, he said, and you will see a different movie every time: infinite possible worlds lost in the tiniest of chances. And yet it is all too tempting to search for general rules, and sometimes we are rewarded. What if we could predict what creatures can survive multiple extinctions, on the scale of half a billion years?
This apparently absurd proposition is what a recent study by Matthew L. Knope, Jonathan L. Payne et al. has shown at the end of this year’s February. They analyze a huge amount of living and fossil marine genera -30,074 and 19,992 respectively, another proof of the power of big data when dealing with the history of life- and try to correlate the taxonomic diversity to the ecological diversity over time. That is, the number of species of a clade versus the ecological strategies exploited by members of that clade.
One could find it obvious that there is some correlation -after all, if you have lots of genera, you can imagine there are more chances they evolved to different niches- but it is not necessarily always so. And this is the main finding of Knope et al.: this correlation evolves over geological time.
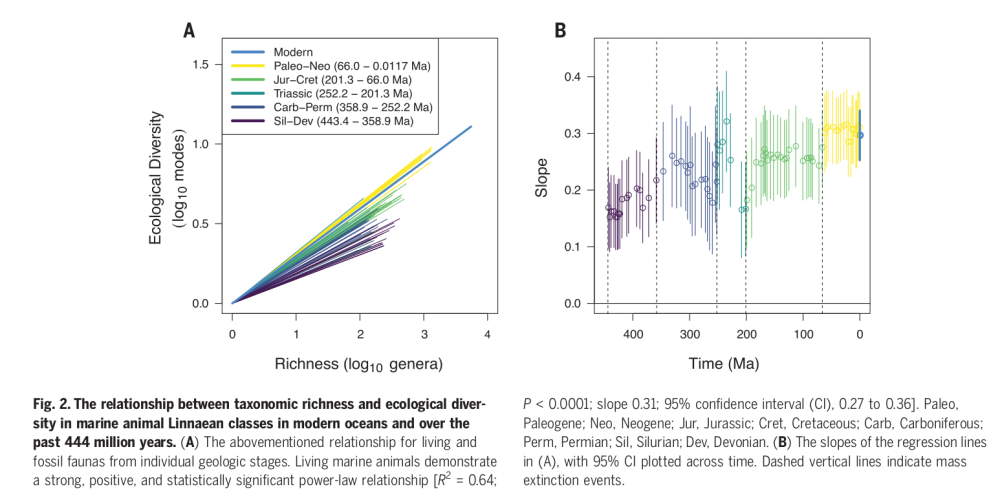
The graph on the left shows the (log-log) correlations between the richness of genera on the x axis and the ecological diversity on the y axis, divided by geological period. The “rays” must be interpreted dynamically. The dark lines laying low are the correlation as it were in the Paleozoic, starting from the Silurian and Devonian. Going forward the correlation shifts: turquoise and green lines, corresponding to the Mesozoic, grow steeper, until in the Tertiary -yellow lines- it becomes almost double than the Paleozoic one. In other words, the more you go forward in time, the more ecological diversity correlates with taxonomic diversity.
Now, this is nice but hard to interpret. Now see the graph on the right: the slopes of the lines on the left are plotted against time. You can see the vertical dashed lines separating mass extinctions. It is not always convincing, but there is some hint that mass extinctions did something. After each extinction, ecological diversity becomes more tightly related to taxonomic ones.
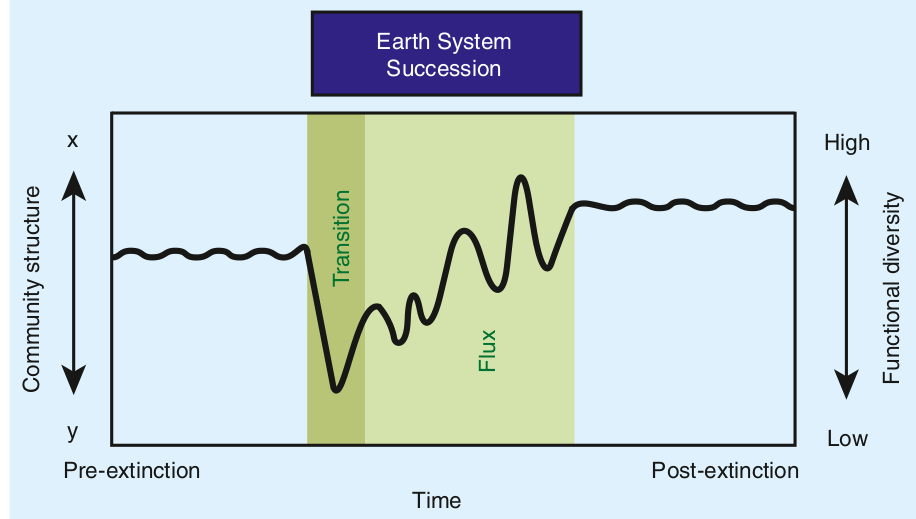
What is going on? We are seeing how extinctions shape the composition of life on the scale of the Phanerozoic. It is not that ecologically diverse clades are better at diversifying and evolving: actually what they find is that, at least in the Tertiary, they diversify less than ecologically poor clades. Each ecological crisis instead tends to wipe out clades that were not flexible enough, forced to live within a few ecological modes. Clades that instead could exploit more ecological niches were able to find at least some of their members surviving a crisis. They diversify and repopulate the oceans for the simple reason they are the ones surviving, and they survive because they have the strategies to do so.
The classes that had high taxonomic richness and low ecological differentiation during the Paleozoic, such as rhychonelliform brachiopods and crinoids, consisted largely or entirely of nonmotile suspension feeders that mostly cannot occupy infaunal or pelagic habitat tiers. In contrast, the classes and phyla that are genus-rich in the Cenozoic (e.g., mollusks, arthropods, and vertebrates) are generally motile, feed in a variety of ways, live across many habitat tiers, have more control over gas exchange with the environment, and have weathered mass extinctions well.
Let’s take it in what this means. First, this means that we have some hope of predicting what animals groups would survive extinction better than others. Animals who are more motile, more versatile, that exploit more environments and more strategies will have better changes. Brachiopods were dominant in the Paleozoic, but a glance at their physiology compared to that of bivalves, in hindsight, shows us they didn’t have a chance. They were simply too rigidly adapted to a single mode of life to fare well after a catastrophe such as the end-Permian.
Second: Our seas are not different from the ones of the Paleozoic just because we have different animals. Our ecosystems have been selected. The animals we have today in the sea are quicker, more mobile, smarter at exploiting different niches, have better metabolisms – and they are like that because they are the hardened survivors of multiple ecological crises. As Knope explains:
The oceans we see today are filled with a dizzying array of species in groups like fishes, arthropods, and mollusks, not because they had higher origination rates than groups that are less common, but because they had lower extinction rates over very long intervals of time.
We are seeing natural selection acting not on the scale of the individual or of the species: we are seeing it acting on entire ecosystems on the scale of half a billion years. Knope et al. show that the shadow of Darwin is longer than we imagined, as long as the tape of Gould, shaping not only single species but the entire composition of life along hundreds of millions of years.
Paper: Knope et al., “Ecologically diverse clades dominate the oceans via extinction resistance“, Science 367, 1035–1038 (2020)