In a previous post on this lethargic blog, I briefly touched the Antarctic extinction, the mass extinction of the fauna and flora inhabiting a whole continent, reduced to a desert of ice.
As is too often happens, I worried myself only about the macroscopic biota, and lazily forgot the microscopic lifeforms. Luckly, scientists are smarter than I am. Eveline Pinseel and coworkers have now described, in a paper on Science Advances, what happened to some of the most iconic microbial taxons – diatoms – that inhabited Antarctica (I have to thank Sylvie Coyaud for bringing this to my attention). Diatoms are special for many reasons, but mainly as being an example of microorganism with incredibly beautiful and unique shells, that readily fossilize and can be classified into prehistoric species and genera, something that it is hardly possible with many other unicellular beings (a notable exception are foraminiferans).

The study shows that in the Miocene (14-15 millions of years ago) Antarctic lakes had a rich diatom flora, with unique species so far unknown to science, but strongly related to those now present in New Zealand, Tasmania; and was overall not unlike the flora present in the contemporary Arctic. Temperatures, now hovering around -12° C, were around +5 °C at the time.
Then, the ice came. In the Pleistocene, about 150.000 years ago, the climate of Antarctica was much similar to today and, correspondingly, diatoms were much less diverse than in the Miocene, but still more diverse than today. The last glacial period was the final hammer for the Antarctic diatom flora: more diatom species were wiped out, not unlike the mammoths, or were confined to sub-Antarctic realms. The diatoms of today’s Antarctica land are a relict of what was once a diverse flora, adapted to some of the harshest conditions on Earth.
According to the authors
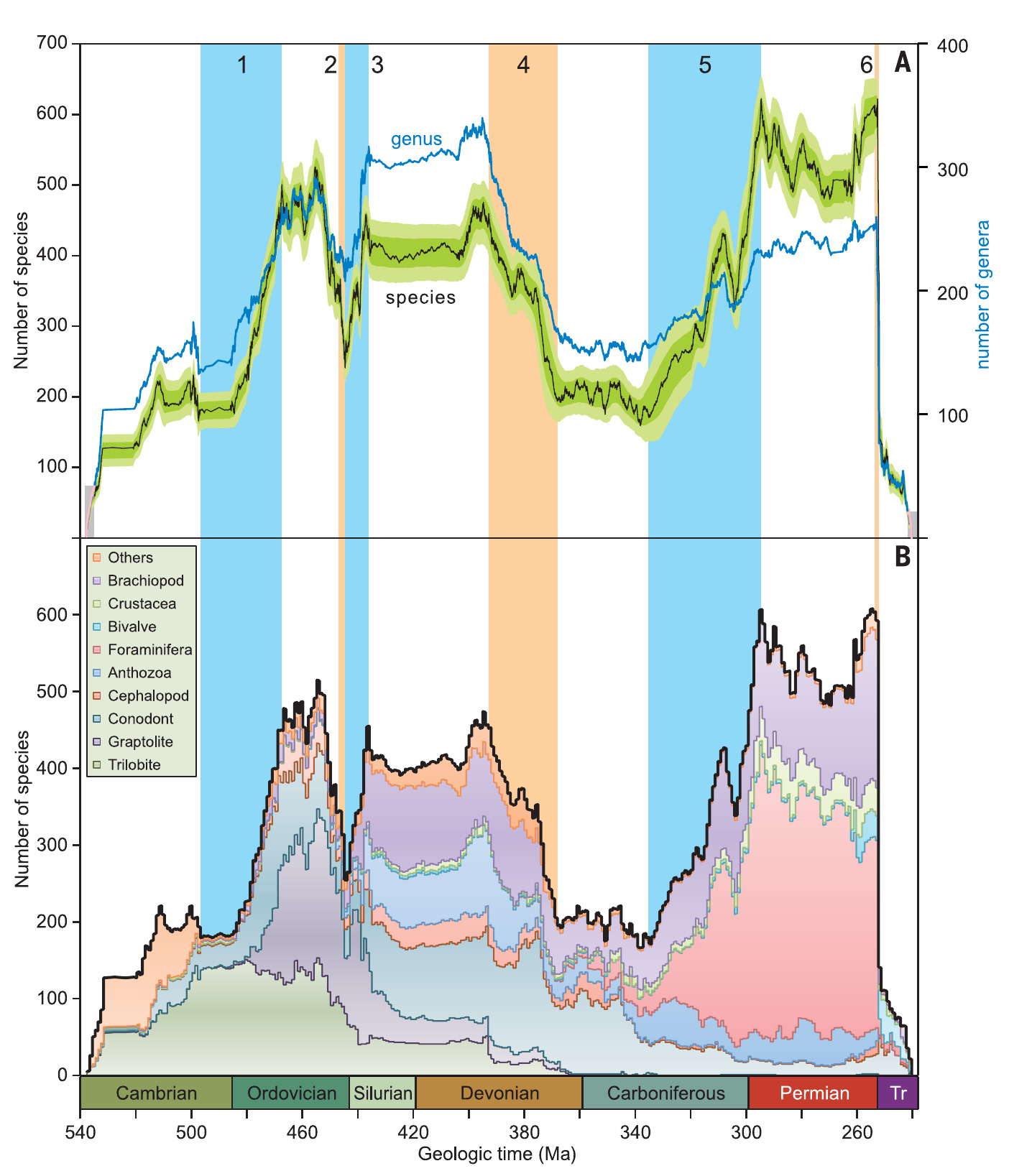
Although there are multiple records of regional extinctions and species turnover of lacustrine diatoms in Quaternary paleorecords of the Northern (60) and Southern (61) Hemisphere, the scale of the extinction of diatoms in continental Antarctica since the mid-Miocene is, both at the species- and genus-level, beyond anything reported in the literature thus far.
In other words, the Antarctic extinction was also the most profound mass extinction of diatoms known stodayo far. Their fate was most probably shared by large parts of the Antarctic microflora, of which we cannot say anything only because they didn’t leave shells to fossilize. Mass extinctions ,therefore, events reshape biodiversity at all levels. This is not news per se, we know that for example the end-Cretaceous extinction led to the extinction of numerous foraminiferan species. But it is a somber reminder of how much biodiversity has been lost forever, how much is going lost now, without us even noticing, without us being even able to interpret what such a loss would mean for other lifeforms.
The paper is: Eveline Pinseel et al. “Extinction of austral diatoms in response to large-scale climate dynamics in Antarctica”, Science Advances, Volume 7, Issue 38, 15 September 2021, https://doi.org/10.1126/sciadv.abh3233